
Gabriel Travis Laboratory
Visual Cycle and Inherited Photoreceptor Degenerations

Gabriel H. Travis, M.D.
Professor
Charles Kenneth Feldman Chair in Ophthalmology
Department of Ophthalmology and Biological Chemistry
Jules Stein Eye Institute
Dr. Travis has been working in the field of vision science since 1989, when he identified the gene affected in the spontaneous retinal degeneration slow (rds) mutant mouse as a postdoctoral fellow. This was one of the first genes causing neural degeneration to be identified. Since then, Dr. Travis has been working on several retinal proteins required for visual function in vertebrates. His approach has been to determine what each protein 'does for a living' normally, and to understand how loss of the protein causes pathology in genetically modified mice or zebrafish, and in patients with mutations in its gene. Methodologies commonly used by his group include molecular biology, biochemistry, analysis of visual retinoids, immunocytochemistry of retina sections, and photoreceptor physiology, in collaboration with Sampath at Jules Stein Eye Institute. The project described below represents one of several currently ongoing in Dr. Travis laboratory.
Characterization of the alternate visual cycle for regeneration of cone-opsin pigments under daylight conditions.
Vertebrates, including humans, use bleaching visual pigments that dissociate following photoisomerization to yield free all‑trans‑retinaldehyde (atRAL) and unliganded apo-opsin. Apo-opsins regain light sensitivity by recombining with an 11‑cis‑retinaldehyde (11cRAL) chromophore molecule. Rhodopsin visual pigment in rods has long been known to utilize the canonical visual cycle in retinal pigment epithelial (RPE) cells. This enzyme pathway is active in the dark and keeps up with rhodopsin photoisomerization under dim light. However, the throughput of this pathway much slower than the rate of visual-pigment photoisomerization in a daylight-exposed retina. A second pathway in Müller glial cells has a higher throughput that keeps up with rhodopsin and cone-opsin photoisomerization in bright daylight. Dr. Travis' group showed that this pathway utilizes RDH10, a retinol dehydrogenase in Müller cells that oxidizes all‑trans‑retinol (atROL) to atRAL, and reduces 11cRAL to 11‑cis‑retinol (11cROL). This enzyme works in conjunction with RGR-opsin, a 'reverse' photoisomerase that converts atRAL to 11cRAL upon exposure to light. More recently, his group showed that RDH12 reduces atRAL to atROL, and oxidizes 11cROL to 11cRAL. Importantly, the Rdh12 gene is affected in infants and children with the severe blinding disease, Leber Congenital Amaurosis. A cartoon of the alternate visual cycle is shown in Figure 1. Because this pathway is driven by light, its throughput scales with light intensity. The energy of chromophore regeneration comes from sunlight, unlike the canonical visual cycle in RPE cells that requires metabolic energy. While rods can only regenerate photobleached rhodopsin with 11cRAL chromophore, cones can use 11cROL (chromophore precursor) or 11cRAL. The alternate visual cycle in Müller cells releases 11cROL, which is taken up and converted by RDH12 in cones to 11cRAL, which they use to regenerate photobleached cone-opsins (Figure 1). Thus, the alternate visual cycle acts as a 'private pipeline' of chromophore precursor that exclusively regenerates cone visual pigments and helps cones to escape competition from rods for scarce 11cRAL chromophore in a daylight illuminated retina.
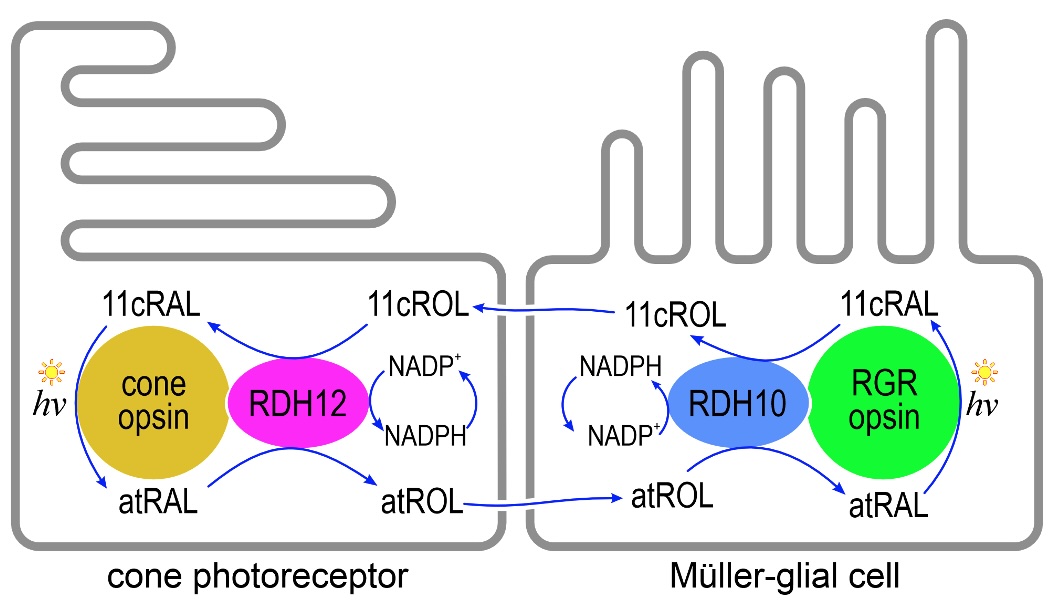
Figure 1. Proposed model for the photic cone visual-cycle in Müller glial cells and cone photoreceptors. Absorption of a photon (hv) by a cone-opsin pigment isomerizes the 11cRAL to atRAL, activating visual transduction. Shortly afterwards, atRAL dissociates from the opsin, is reduced to atROL by ZCRDH and is released into the extracellular space for recycling. The atROL is taken up by the Müller cell where it is re-oxidized by RDH10 to atRAL, which binds covalently to apo-RGR opsin. Absorption of a photon by RGR opsin isomerizes atRAL to 11cRAL. The 11cRAL is reduced, again by RDH10, to 11cROL, which is released by Müller cells into the extracellular space. Finally, the 11cROL is taken up by the cone photoreceptor where it is oxidized by RDH12 to 11cRAL, which combines with cone apo-opsin to form a new cone visual pigment. Since only cones can utilize 11cROL to regenerate their visual pigments, the visual cycle in Müller cells helps cones avoid competition with rods for chromophore under bright-light conditions.
Recent Publications
Dual Knockout Models of the Spatially and Functionally Conserved rgra and rgrb Zebrafish Genes Reveal the Requirement of RGR for the Integrity of Cone‐Mediated Photopic Vision, the Photopic Visual Cycle and Bruch's Membrane Morphology
Grace Ruddin, Tess McCann, Joanna J. Kaylor, Michelle M. Fox, John D. Fehilly, Rebecca Ward, Adam Faulkner, Ailís L. Moran, Kieran Wynne, Roxana A. Radu, Michael G. Monaghan, Stephen D. Thorpe, Gabriel H. Travis, Breandán N. Kennedy
The FASEB Journal (2026)
View PublicationThe dark and bright sides of retinal G protein-coupled receptor (RGR) in vision and disease
Grace Ruddin, Tess McCann, John D. Fehilly, Jodie Kearney, Breandán N. Kennedy
Progress in Retinal and Eye Research (2025)
View PublicationGermline Disruption of Retinal Pigment Epithelium-Expressed Zebrafish rlbp1b-/- Results in Selective Dim Light Visual Behavior Deficits and Provides a Screening Platform for Evaluating the Pathogenicity of Human RLBP1 Variants
John Denis Fehilly, Tess McCann, Grace Ruddin, Joanna J. Kaylor, Hannah Grenville, Rebecca Ward, Kieran Wynne, Alicia Gómez Sánchez, Elin Strachan, Gabriel Travis, Ross F. Collery, Breandan Kennedy
The FASEB Journal (2025)
View Publication